4.4. Кручення групи рослинних матеріалів
При роботі
сільськогосподарських машин кручення групи матеріалів, хоча і рідко, але має
місце. Така деформація відбувається при роботі перевертача стрічки зі стебел
льону, при повороті групи рослин у бральних рівчаках льонозбиральних машин
внаслідок різних швидкостей
руху бральних пасів; до деякої міри така деформація має місце при пресуванні
стебел у просторі між 4-6 вальцями, що обертаються в одну сторону, при
захопленні і навантаженні сіно-соломистих матеріалів. Перевертач стебел
закручує стрічку з паралельних стебел, при цьому пари сил, що викликають
кручення, діють в площині, паралельній лініям стебел і перпендикулярній площині
стрічки. Поворот групи стебел у бральних рівчаках здійснюється під дією пари
сил у площині, перпендикулярній стеблам.
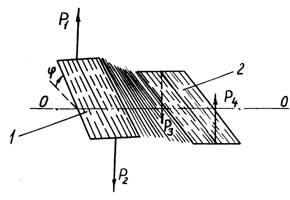
На
рис. 4.46 показана схема стрічки з паралельних рослин, затиснутих на кінцях
затискачами 1 і 2, на які діють протилежно спрямовані пари сил (Р1, Р2) і (Р3, Р4). Під дією цих пар сил одна частина стрічки
повертається щодо іншої навколо осі ОО
на кут j. Слід зазначити, що цілком паралельними стебла будуть
рідко, деякий перекіс у їхньому укладанні можливий.
|
|
Рис. 4.46. Дія на стрічку
з паралельних стебел з метою кручення однієї її частини щодо іншої навколо
осі ОО: 1 і 2 – затискачі |
Під час повороту стрічки
переборюється опір унаслідок зчеплення стебел одне з одним або їхнього
переплетення (у місцях, де стебла перехрещуються, тобто де порушена
паралельність). Зчеплення стебел може мати місце і через їхнє сплетіння плодоніжками
або насінними коробочками.
Ряд
отриманих при експериментальному дослідженні діаграм кручення, що являють собою
залежність моменту Мк пари
сил (Р1, Р2) від кута закручування,
представлений на рис. 4.47, з них на рис. 4.47, а показана діаграма для погано зчеплених стебел, а на рис. 4.47, б – для стебел, зчеплених досить сильно.
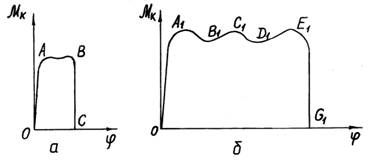
Рис. 4.47. Діаграми кручення шару паралельних
стебел: а – стебла погано зчеплені; б – стебла сильно зчеплені
При крученні погано
зчеплених стебел момент Мк
швидко досягає найбільшого значення (ділянка ОА), потім після деякого коливання (ділянка АВ) відбувається розрив стеблевої стрічки (ділянка ВС); ділянка ОС, що відповідає всьому куту закручування стебел, тут малий, кут
закручування мало залежить від відстані між затискачами 1 і 2 (рис. 4.46),
оскільки розрив стрічки відбувається в найбільш погано зчепленій її частини; до
деякої міри впливає на значення моменту Мк
власна вага стебел, що повертаються.
При крученні сильно
зчеплених стебел (рис. 4.47, б)
момент Мк також швидко
досягає великого значення (ділянка ОА1),
потім після ряду його коливань у бік збільшення або зменшення (ділянка A1B1C1D1E1)
відбувається розрив стеблової маси в найбільш слабкій її частині (ділянка E1G1). Ділянка OG1, що відповідає всьому
куту закручування стебел, тут значно більша, ніж у випадку на рис. 4.47, а, кут закручування тут мало залежить
від відстані між затискачами 1 і 2 (рис. 4.46), тому що розрив стрічки
відбувається в найбільш погано зчепленій її частині.
Кут закручування, що відповідає ділянці OG1
(рис. 4.47, б), залежить від числа рослин на
одиницю довжини стрічки і ступеня їхнього
сплетення (зчеплення). На значення моменту Мк
впливає і власна вага рослин, що повертаються.
Властивість стеблевої
стрічки чинити опір руйнуванню при крученні за схемою на рис. 4.46 цілком може
бути охарактеризовано максимальним значенням Мк max моменту Мк,
необхідним для руйнування стрічки. Аналіз експериментальних даних показав, що
![]() , (4.96)
, (4.96)
де ![]() - дослідний коефіцієнт; iм - число стебел на 1 м довжини
стрічки.
- дослідний коефіцієнт; iм - число стебел на 1 м довжини
стрічки.
Коефіцієнт ![]() залежить від зчеплення
стебел, їхнього діаметра і вологості; знаходиться він для стебел льону, розташованих у стрічці шириною 0,8…1,2 м, у межах
(1...6)×10-5 Н×м2. У сильно розрідженої стрічки, коли стебла мало зчеплені, Мк
max ®0.
залежить від зчеплення
стебел, їхнього діаметра і вологості; знаходиться він для стебел льону, розташованих у стрічці шириною 0,8…1,2 м, у межах
(1...6)×10-5 Н×м2. У сильно розрідженої стрічки, коли стебла мало зчеплені, Мк
max ®0.
При розгляді кручення групи
паралельних стебел навколо осі ОО, їм
паралельної (рис. 4.48), варто виділити окремо центральне стебло, вісь якого
збігається з віссю ОО, і периферійні
стебла, що розташовані довкола нього.
|
|
Рис. 4.48. Кручення групи паралельних стебел |
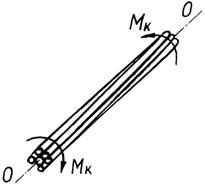
Кручення центрального стебла групи відбувається за
закономірностями, описаними у п. 3.4, а кручення кожного периферійного стебла
навколо центрального складається з ряду вигинів його елементів навколо осей,
положення яких по довжині стебла змінюється. При такому крученні усередині
групи стебел у місцях їхнього дотику один з одним діють ще сили тертя.
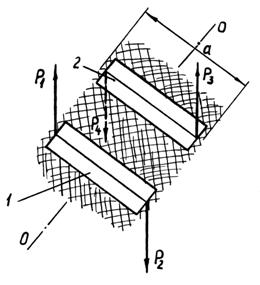
Схема кручення групи безладно вкладених рослин,
положення кожної з яких випадкове, показане на рис. 4.49. Шар рослин затиснутий
у двох місцях затискачами 1 і 2, на які діють пари сил (Р1, Р2) і (Р3, Р4),
спрямовані в протилежні сторони. Під дією цих сил відбувається поворот однієї
частини шару рослин відносно іншої навколо осі ОО на кут, який позначимо j. Під час повороту
переборюється опір унаслідок сплетіння рослин, зчеплення і тертя між ними.
|
|
Рис. 4.49. Вплив на шар рослин з метою кручення
однієї його частини відносно іншої навколо осі ОО: 1 і 2 - затискачі |
Оскільки розташування рослин
у шарі випадкове, то і діаграма кручення таких рослин носить випадковий
характер; вона може мати такі вигляди, як представлені на рис. 4.47, але може
мати й інші вигляди. Опір такого шару стебел руйнуванню при крученні також може
бути охарактеризований максимальним значенням моменту Мкmax, при якому відбувається руйнування шару рослин.
Момент цей залежить від висоти шару h,
його ширини a (рис. 4.49), щільності
стеблової маси і відстані між затискачами. Наближено він визначається за
формулою (при a, рівному 0,5…1,5м, і h < 1,0 м):
![]() , (4.97)
, (4.97)
де ![]() - дослідний коефіцієнт; r - щільність стеблової маси; b - відстань між затискачами.
- дослідний коефіцієнт; r - щільність стеблової маси; b - відстань між затискачами.
При а = 0,8...1,4м і h <
1,0 м к0 знаходиться в межах 0,02...0,08 м5/с2.
Дослідження явища післядії
при крученні групи стебел проводиться на приладі, представленому на рис. 4.50,
але може проводиться і на інших приладах. За результатами дослідів установлено,
що закономірності при релаксації напружень і I стадії повзучості у скручених
груп стебел принципово такі ж, які
представлені на рис. 3.40, а і б та які відповідають рис. 3.9.
Справедливі також для цього випадку й отримані раніше залежності (3.40),
(3.41), (3.42), (3.43), (3.44), (3.45), (3.46), (3.47) і
|
|
Рис. 4.50. Прилад для кручення стебел із встановленими в ньому стеблами льону для дослідження релаксації напружень |

(3.48), що описують падіння моментів і ріст кутової
деформації в рослин, які скручуються.
Обробкою за
наведеними формулами результатів дослідів з кручення груп стебел встановлено,
що релаксація моментів при крученні групи стебел жита вологістю 15% і середнього діаметра 3,0 мм мала місце від моменту Мк0, рівного 0,032 Н×м, до моменту Мкк,
рівного 0,022 Н×м, час tк
був рівний 3,0 годинам, ![]() було – 0,06 Н×м×год-1, а показник
χр дорівнює 1,46. При крученні групи стебел
льону вологістю 15 % і середнім діаметром 1,2 мм релаксація моментів мала
місце від Мк0, рівного
0,145 Н×м, до Мкк,
було – 0,06 Н×м×год-1, а показник
χр дорівнює 1,46. При крученні групи стебел
льону вологістю 15 % і середнім діаметром 1,2 мм релаксація моментів мала
місце від Мк0, рівного
0,145 Н×м, до Мкк,
рівного 0,130 Н×м, час tк
був рівний 2,0 годинам, ![]() було -0,010 Н×м×год-1, а показник χр дорівнював 1,12. Кручення таких же стебел при
інших значеннях моментів привело до їхньої релаксації від Мк0, рівного 0,110 Н×м, до Мкк,
рівного 0,095 Н×м, час tк
дорівнював 2,0 години,
було -0,010 Н×м×год-1, а показник χр дорівнював 1,12. Кручення таких же стебел при
інших значеннях моментів привело до їхньої релаксації від Мк0, рівного 0,110 Н×м, до Мкк,
рівного 0,095 Н×м, час tк
дорівнював 2,0 години, ![]() було -0,015 Н×м×год-1, а показник χр
був рівним 1,16.
було -0,015 Н×м×год-1, а показник χр
був рівним 1,16.

