Тема 18.
Водорості: особливості будови, екологічні групи, систематика
Водорості – Аlgae – це нижчі таломні, або сланеві спорові рослини, що містять у
своїх клітинах фотосинтезуючі пігменти і живуть переважно у воді.
Основною структурною одиницею талому водоростей
є клітини. Вони можуть бути голі або вкриті різними покривами – пектиновою чи
пектиново-целюлозною оболонкою, кремнеземовою текою або іншими мінералізованими
покривами.
Протопласт водоростей, за винятком
прокаріот, диференційований на цитоплазму з органоїдами і ядро. Клітини
переважно одноядерні, але бувають дво-, три- і багатоядерні. Серед органоїдів
найбільшої уваги заслуговує хлоропласт, що складається з двомембранної
оболонки, строми і ламел, що не утворюють гран. У хлоропластах є піреноїди –
білкові тільця, що синтезують полісахариди.
Водорості, за винятком червоних,
синьозелених і деяких зелених, рухаються самі або утворюють рухомі стадії.
Здатність до руху забезпечується джгутиками, війками або несправжніми війками.
Талом водоростей – одноклітинний,
колоніальний, неклітинний або багатоклітинний. Вони утворюють дев’ять основних
типів морфологічної структури талому: амебоїдну, монадну, кокоїдну,
пальмелоїдну, нитчасту, різнонитчасту, пластинчасту, сифональну і харофітну.
Розмноження водоростей – вегетативне,
безстатеве і статеве. Вегетативно розмножуються частками слані або спеціальними
бруньками, бульбочками, акінетами; безстатево – зооспорами або спорами.
Статеве розмноження дуже різноманітне: хологамія,
автогамія, кон’югація, ізогамія, гетерогамія, оогамія. У багатьох водоростей має
місце чергування спорофіта і гаметофіта.
За пристосуванням до різноманітних умов
водорості діляться на такі екологічні групи: водні (планктонні, бентосні,
перифітонні), аерофітні, ґрунтові, водорості гарячих джерел, водорості снігу і льоду, водорості
солоних водойм, водорості вапнякових субстратів.
До водоростей належить близько 40 тис.
видів, які ділять на десять (шістнадцять) відділів, переважно за забарвленням
і, особливостями будови: синьо-зелені, динофітові, золотисті, діатомові,
жовто-зелені, бурі, червоні, евгленові, зелені.
Сучасна
систематика водоростей
Відділ
Синьозелені водорості – Cyanophyta
Відділ
Евгленофітові водорості – Euglenophyta
Відділ
Хлорарахніофітові водорості – Chlorarachniophyta
Відділ
Рафідофітові водорості – Raphidophyta
§ Порядок
Raphidiales (Vacuolariales)
Відділ
Золотисті водорості – Chrysophyta
Відділ
Евстигматофітові водорості – Eustigmatophyta
Відділ
Жовтозелені водорості – Xantophyta
Відділ
Бурі водорості – Phaeophyta
Відділ
Діатомові водорості – Bacillariophyta
o Клас
Coscinodiscophyceae (Centrophyceae)
·
Відділ
Диктіохофітові водорості – Dictyochophyta
Відділ
Жовто-зелені водорості – Xantophyta
Це
одноклітинні, колоніальні або багатоклітинні водорості світло-, темно-жовтого
або жовто-зеленого кольору, дуже рідко без кольору. Клітинна оболонка
пектинова, з невеликою домішкою целюлози, часто мінералізована. Ядро
здебільшого одне, хроматофори зернисті, іноді з піреноїдами, містять хлорофіл а,
с, е, ксантофіл і каротиноїди. Запасшіми речовинами є олія, лейкозин,
волютин, хризоламінарин; крохмаль ніколи не утворюється.

Рис. 18.1. Жовтозелені
водорості
Вегетативне
розмноження здійснюється шляхом поділу клітин або частинами талому. Статевий
процес не характерний для жовто-зелених і зустрічається порівняно рідко.
Поширені жовто-зелені водорості у водоймах, з прісною, рідше солоною водою, де
вони є важливим компонентом планктоку. а іноді й бентосу, оселяються у грунті,
на каменях.
До відділу
Жовто-зелені водорості належить вошерії. Вошерія утворює в природних умовах
скупчення ниткоподібних тенно-зелених таломів у прісній стоячій і проточній
воді, а також по берегах висихаючих водойм, прикріпляючись до грунту за
допомогою ризоїдів. Талом неклітинний, гіллястий, багатоядерний, з дрібними
хроматофорами. Клітина містить велику кількість ядер.
Відділ
синьо-зелені водорості – Сyаnорhytа
Синьозелені водорості – це прокаріотні одноклітинні, колоніальні
або багатоклітинні рослини. Структура талому кокоїдна, пальмелоїдна або
нитчаста. Клітина вкрита оболонкою, утвореною пектиновими речовинами,
слизистими полісахаридами і целюлозою. Вона ослизнюється або утворює спеціальну
слизову піхву.
Протопласт не диференційований на ядро і
цитоплазму з органоїдами. Він поділяється на хроматоплазму – зовнішню, щільну,
інтенсивно забарвлену і центроплазму – внутрішню рідку слабозабарвлену.
При детальному вивченні клітини було
виділено три її частини: нуклеоплазму, фотосинтезуючі ламели, рибосоми та інші
цитоплазматичні гранули. В нуклеоплазмі міститься ДНК, а у складі
фотосинтезуючих ламел – хлорофіл а, каротиноїди, фікоціан і фікоеритрин.
Продукти фотосинтезу – глікопротеїди, полісахариди, волютин.
Нитчасті форми синьо-зелених водоростей
можуть утворювати колонії або гормогоніальну структуру. Для гормогонієвих
характерні своєрідні клітини –гетероцисти. Це безбарвні клітини з подвійною
оболонкою. Вони не мають газових вакуолей і не містять запасних поживних
речовин. Ділянки із забарвлених клітин між сусідніми гетероцистами називаються гормогоніями.
За способом живлення – це автотрофічні
рослини, але при відповідних умовах вони можуть переходити на гетеротрофне
живлення. Таке змішане живлення називається міксотрофним.
Розмножуються синьо-зелені водорості поділом клітин
(одноклітинні) або
гормогоніями (нитчасті). Статевий процес відсутній.

Рис. 18.2. Синьозелена водорість носток:
а –загальний вигляд; б – вигляд при малому збільшенні мікроскопа; в – вигляд
при великому збільшенні мікроскопа: 1 – гормогоній; 2 – гетероциста
Синьозелені
водорості відзначаються широкою амплітудою екологічного пристосування. Вони
зустрічаються при температурах від плюс 75 °С до мінус 83 °С. Більшість із них
планктоні розвиваються і викликають „цвітіння” води. Поселяються водорості у
ґрунті і на ґрунті, на корі дерев, скелях, у сланях лишайників тощо.
Відділ синьозелених водоростей поділяють
на три класи: хроококові, хамесифонові, гормогонієві.
Відділ
Зелені водорості – СНLORОРНYТА
Зелені водорості - це одноклітинні,
колоніальні, неклітинні або багатоклітинні нижчі рослини. Талом їх має різні
типи морфологічної структури: монадну, кокоїдну, пальмелоїдну, нитчасту,
пластинчасту, сифональну. Клітини вкриті пектиновою, пектиново-целюлозною або
целюлозною оболонкою подібно до вищих рослин. Протопласт диференційований на
цитоплазму з органоїдами і ядро. Серед органоїдів цитоплазми найважливішим є
хлоропласт з піреноїдами. У його ламелах містяться такі самі пігменти, як і у
вищих рослин – хлорофіли а і в, а також специфічні для водоростей
– c і d, каротин і ксантофіли. Запасним полісахаридом, як і у вищих рослин,
здебільшого є крохмаль.
Розмноження вегетативне (частинами слані
або бульбочками), безстатеве (дво- або чотириджгутиковими зооспорами чи
автоспорами) і статеве (хологамія, кон’югація, ізогамія, гетерогамія, оогамія).
Поширені переважно в прісних водоймах,
але є солоно-водні, а також ґрунтові і наземні аерофітні водорості.
Цей відділ
об'єднує одноклітинні, колоніальні, багатоклітинні організми. Клітини зелених
водоростей можуть бути одно або багатоядерними; клітинна оболонка
целюлозо-пектинова, але в деяких представників оболонка відсутня і клітини
покриті лише цитоплазматичною мембраною. Клітини мають зелений колір,
зумовлений наявністю хлорофілу а і в; крім того, тут є каротин,
ксантофіли, У хроматофорах розміщені піреноїди, навколо яких відкладається
запасний крохмаль і олія.
Зелені
водорості поділяються на три класи: справжні зелені, кон'югати, харові.

Рис.
18.3. Різноманітність зелених водоростей
Клас
Справжні зелені, або Рівноджгутикові водорості
Типовими
представником цього класу є хламідомонада, а також хлоpeлa, улотрикс та
плеврокок.
Хламідомонада
– одноклітинна мікроскопічна водорость, яка розмножується статевим і нестатевим
способом (рис. 17.1). При нестатевому розмноженні клітина втрачає джтутики, ділиться її
ядро, хроматофор і цитоплазма на 4 (рідше 8) клітин – зооспор. У кожної
дочірньої клітини виростають по 2 джгутики, оболонка материнської клітини
руйнується і зооспори виходять у воду. Таким чином водорості розмножуються дуже
швидко. Вже через добу дочірні клітини знов діляться.
При статевому
розмноженні у материнській клітині утворюються гамети. Вони подібні до зооспор,
але їхня кількість значно більша: 32 або 64 в одній клітині. Після дозрівання
гамети виходять із материнської клітини і попарно сполучаються, утворюючи
зиготу. Вона вкривається захисною оболонкою і переходить у стан спокою. Через
деякий час зигота виходить і:5 оболонки і ділиться мейотично з утворенням 4
гаплоїдних зооспор.
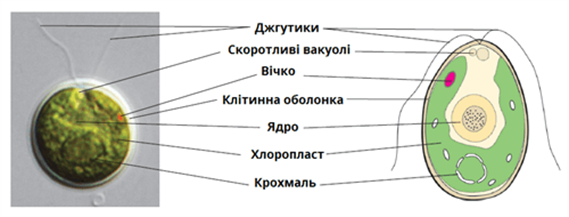
Рис. 18.4.
Будова хламідомонади (Chlamydomonas sp.)
Клас Кон'югати
представлений одноклітинними,
колоніальними і багатоклітинними організмами. Клітини одноядерні, з
хроматофором зірчастої, пластинчастої або стрічкоподібної форми. Розмножуються
кон'югати поділом клітин і статево - кон'югацією. Нестатевого розмноження немає.
Спірогіра – типовий представник кон'югат. Її види
живуть у прісних водоймах – річках, ставках, озерах і торф'яних болотах. Довгі
нитки талому утворюють сплетення (баговиння) яскраво-зеленого кольору, до
субстрату вони не прикріплюються і вільно плавають у воді. Оболонка клітини –
целюлозно-пектинова, без пop. Цитоплазма займає пристінне положення. В
пристінному шарі цитоплазми знаходяться один або кілька спірально закручених
стрічкоподібних хроматофорів. Хроматофори з піреноїдами, по краю більш-менш розсічені,
ядро міститься у центрі, вакуолей в клітині кілька.
Розмножується спірогіра вегетативно і статево.
Вегетативне розмноження відбувається під час випадкового розриву ниток або
розпаду на окремі нитки за несприятливих умов. Декілька клітин ослизнюються і
відмирають. Такі мертві клітини називаються гетероцистами. У місці виникнення
гетероцисти нитка спірогіри розпадається на дві. З кожної частини нитки
утворюються нові особини за рахунок поділу клітин.
Статевий процес – кон'югація – відбувається так; дві
нитки, зовні подібні, зближуються, в їхніх клітинах виникає випинання стінок,
що ростуть одна одній назустріч. Під час контакту цих виростів перегородки їх
зникають і вміст "чоловічої" клітини переливається в
"жіночу". В результаті цього злиття утворюється зигота, яка
вкривається товстою оболонкою і після певного періоду спокою ділиться
мейотично, при цьому утворюється 4 гаплоїдних клітини, три з них відмирають, а
одна проростає і дає початок новій особині. Таким чином, у кон'югатів у
життєвому циклі переважає гаплоїдна фаза, диплоїдна лише зигота.
|
|
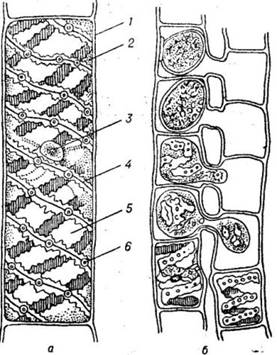
Рис. 18.5. Зелена водорість спірогіри:
а – будова клітини спірогіри; б –
кон'югація спірогіри: 1 – клітинна оболонка; 2 – цитоплазма; 3 – ядро; 4 –
хроматофор; 5 – вакуоля; 6 – піреноїди |
Клас Харові водорості
Цей клас водоростей
відрізняється від інших зелених водоростей складною будовою макроскопічного,
багатоклітинного талому, що зовні нагадує вищі рослини. Талом у харових
розчленований на стеблову частину, кільчасто розміщені бічні гілки-листки і
розгалужені ризоїди. Ріст талому верхівковий та інтеркалярний. Клітини вкриті
товстою оболонкою, одно- і багатоядерні, хроматофори зернисті, численні.
Розмно-жуються харові вегетативно (частинами слані і бульбочками на ризоїдах) і
статево. Статевий процес оогамний: оогонії і антеридії багатоклітинні.
Розповсюджені харові найчастіше у водоймах з прісною водою. Найпоширенішими є
хара і нітела.
Відділ Діатомові водорості –
Bacillariophyta
До діатомових водоростей
відносяться одноклітинні або колоніальні організми мікроскопічно малих розмірів
з кокоїдною структурою талому. Вони оселяються у солоних і прісних водоймах, на
вологому грунті, скелях, корі стовбурів дерев тощо. Клітини діатомових покриті
панцирем із кремнезему, який складається із двох часток – тек. Більша частка
панцира (епітека) покриває меншу (гіпотеку). В свою чергу, кожна з часток має
плоску частину – стулку з трохи загнутими краями і поясок – вузьке кільце,
щільно з'єднане із стулкою. Цитоплазма з центральною вакуолею займає пристінне
положення. Хроматофори зернисті або пластинчасті з одним чи кількома голими
піреноїдамі, мають жовте або буре забарвлення, зумовлене наявністю, крім
хлорофілу а і с, пігментів фукоксантину і діатоксантину. Запасні
продукти відкладаються у вигляді жирної олії, а також волютину й лейкозину.

Рис. 18.6. Різноманітність діатомових водоростей
Вегетативне розмноження
відбувається шляхом поділу клітини. Потім відбувається розходження і добудова
кожним протопластом нової гіпотеки. Серія таких поділів призводить до
послідовного здрібнення особин. Під час статевого процесу спостерігається
відновлення нормальних розмірів здрібнілих особин. Дві клітини наближаються
одна до одної, скидають теки і обволікаються слизом. Кожна з клітин ділиться
мейотично, внаслідок чого утворюється чотири гаплоїдні клітини - і-тетрада. Дві
клітини різних тетрад зливаються, а решта відмирає. З зиготи виростає нова
особина нормальних розмірів. У життєвому циклі діатомових водоростей переважає
диплоїдна фаза. Залежно від форми і будови стулок діатомові поділяють на два
класи: Центричні та Пенатні.
До класу Центричні відносять
одноклітинні і колоніальні водорості. Клітини цих водоростей характеризуються
радіальною симетріоо, стулки не мають шва, і тому вони не здатні до активного
руху. Поширені центричні в планктоні, бентосі та перифітоні морів і океанів.
Типовими є циклотела, мелозира, ризосоленія.
Клас Пенатні містить
одноклітинні та колоніальні водорості з характерною двобічною симетрією клітин.
Пенатні поширені в прісноводному і морському бентосі, рідше планктоні. Багато
видів зустрічається в грунті. Найлоширенішими є одноклітинні: пінулярія,
плевросигма, навікула, гомфонема і колоніальні – табелярія, діатома, фрагілярія
(рис. 2).
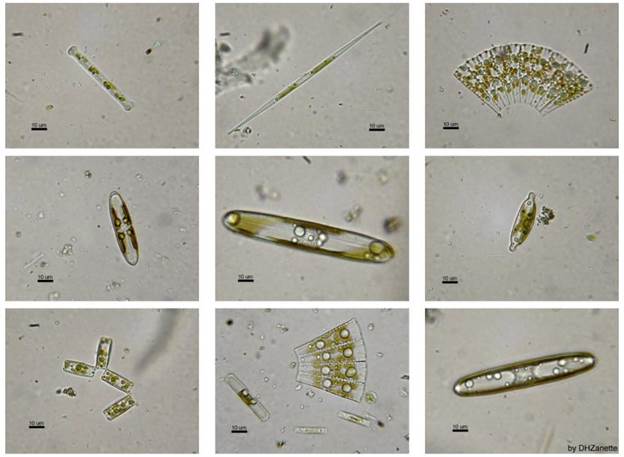
Рис. 18.7. Прісноводні види діатомових
водоростей
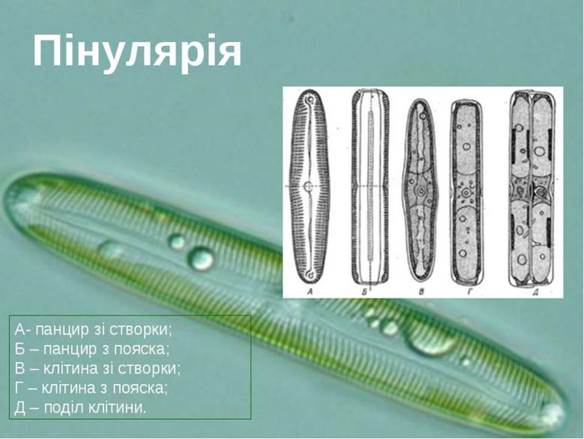
Рис. 18.8. Пінулярія
Відділ Бурі водорості -
Phaeophyta
До цього віддіту
відносяться найбільш високоорганізовані багатоклітинні організми. Ріст талому
верхівковий або вставний (інтеркахіярний). Клітинна оболонка бурих двошарова:
внутрішній шар її целюлозний, зовнішній – пектиновий, часто ослизнений.
Цитоплазма пристінна, з одним ядром і численними дрібними вакуолями. Буре
забарвлення зумовлене наявністю специфічного бурого пігменту фукоксантину, є
також хлорофіли, каротиноїди та ксантофіли. Запасними продуктами є ламінарин,
маніт (цукроспирт) і жирна олія. В клітинах міститься дуже велика кількість
калію.
Розмноження бурих водоростей
здійснюється вегетативно – частинами талому, нестатеве – за допомогою численних
джгутикових зооспор, які утворюються в одноклітинних, зрідка багатоклітинних
зооспорангіях, або нерухомих тетраспор, що утворюються в тетраспорангіях.
Статевий процес ізо-, гетеро-
і оогамний. Ізо- і гетерогамети формуються в багатоклітинних гаметангіях;
оогонії і антеридії одноклітинні. Для більшості бурїіх водоростей характерна
зміна поколінь і ядерних фаз.
Найбільш відомий представник
– ламінарія цукрова або морська капуста. Використовується в їжу у свіжому та
консервованому вигляді. В оболонках клітин містяться альгінові кислоти, які
з'єднуються з радіонуклідами і виводять їх з організму. Великий вміст калію
також робить її корисною при серцевих захворювашшх. Ламінарія пальчаста
використовується в фармакології. З неї виробляють ліки, які містять калій, та
використовують як природні мінеральні добрива (заорюють таломи в грунт). На
таломі водорості саргасум утворюються повітряні міхури. Вони тягнуть
талом до поверхні, ризоїди відриваються від дна і водорості течією Гольфстрім
переносяться до Саргасового моря, де утворюють баговиння, яке затримує
надходження сонячного світла в глибини моря. Тому Саргасове море називають
мертвим. Ці водорості ускладнюють судноплавство.
Відділ Червоні водорості – Rhodophyta
Це переважно багатоклітинні
водорості нитчастої, пластинчастої, паренхімної структур, рідше одноклітинні та
колоніальні, мікроскопічні – кокоїдної структури: рожевого, червоного,
оранжевого, жовтого, фіолетового або майже чорного кольору. Талом має вигляд кущиків,
які складаються з багатоклітинних гіллястих ниток до 2 м завдовжки. Клітинні
оболонки целлозно-пектинові, містять агар і агароїди, тому деякі види
використовують як сировину для добування агару. В інших видів клітинні стінки
–інкрустовані вапном, тому вони тверді, як камені. Такі види беруть участь в
утворенні коралових рифів. Цитоплазма досить в'язка, пристінна, ядро дрібне,
одне, рідше їх багато; вакуоля одна, центральна. Рухливих форм у життєвому
циклі немає. Хроматофори – центральні, зірчасті або пристінні, з піреноїдами;
пластинчасті лінзо- або стрічкоподібні, без піреноїдів, містять специфічний
набір пігментів: хлорофіли а і b, каротин, ксантофіли і фікобіліни –
фікоціанін

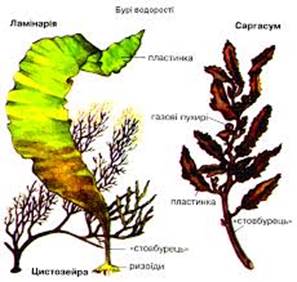

Рис. 18.9. Бурі та червоні водорості
(синього кольору) і фікоеритрин (червоного кольору).
Продуктами асиміляції червоних водоростей є багрянковий крохмаль, який
забарвлюється в червоний колір під дією йоду. Містять велику кількість йоду.
Найпримітивніші представники
червоних водоростей розмножуються виключно вегетативно – шляхом поділу клітин.
Під час нестатевого розмноження на таломі утворюються
спорангії, які містять по одній спорі (моноспори) або по чотири спори
(тетраспори).Статевий процес оогамний.
Представники:
порфіра, родименія, анфельція (використовується для добування агар-агару),
калітамніон (в оболонці відкладається вапняк), делесерія, філофора
(використовують як сировину для добування йоду та апіроїдів).
ЛИШАЙНИКИ
Лишайники –
це своєрідна група живих істот, які утворилися внаслідок симбіозу деяких видів
зелених і синьо-зелених водоростей та деяких видів сумчастих грибів і
базидіоміцетів.
Мікологи
вважають, що лишайники виникли з різних груп грибів, які вступили у співжиття з
водоростями, і тому вони мають бути віднесеними до царства грибів. Згідно з
іншою точкою зору лишайники розглядаються як особливий, самостійний єдиний
відділ рослинного світу, що не відноситься ні до нижчих, ні до вищих рослин.
Лишайники –
це організми, в тілі яких об'єднані водорості й гриби, утворюючи комплексний
симбіотичний організм з новими морфологічними, фізіологічними та екологічними
властивостями. Живляться лишайники симбіотично: гіфи гриба забезпечують
водорості водою та мінеральними речовинами, а також захищають водорість від
яскравого освітлення, сильного перегрівання променями сонця і висихання.
Водорості у процесі фотосинтезу синтезують органічні речовини і передають їх
грибам.
Питання про
взаємовідносини гриба і водорості в тілі лишайника до цього часу остаточно не
з'ясовано. Існують гіпотези про паразитизм гриба на водорості, двобічний
взаємозв'язаний паразитизм, паразитизм водорості на тілі гриба і найбільш
поширена гіпотеза взаємовигідного симбіозу, Відомо більше 20 тис. лишайників.
Від інших організмів, у тому числі і від окремих грибів і водоростей, вони відрізняються
за формою, будовою, характером обміну речовин, особливими лишайниковими
кислотами, способами розмноження, повільним ростом (від 1 до 8 MM зарік).
Слань
лишайника складається з переплетених ниток грибниці – гіф – і розташованих між
ними клітин або ниток водоростей. Розрізняють два основних типи мікроскопічної
структури слані лишайників: гомеомерний і гетеромерний. На поперечному зрізі
лишайника гомеомерного типу видно, що слань утворена хаотично переплетеними
гіфами гриба, поміж яких розкидані окремі клітини або нитки водоростей. Ці
лишайники утворюють групу, відому під назвою слизистих лишайників, оскільки
водорості виділяють слиз, в який занурені клітини гриба і водорості. В
лишайнику гетеромерного типу клітини водоростей зосереджені в одному шарі, який
називається гонідіальним шаром, або альгальною зоною. Під ним знаходиться
серцевина, яка складається із пухко розташованих ниток гриба. Зовнішніми шарами
лишайника є щільно зімкнуті грибні гіфи, які називаються корковими шарами. За
допомогою грибних ниток, що відходять від нижньогї кори, лишайник
прикріплюється до субстрату, на якому росте. У деяких лишайників нижня кора
відсутня і він зростається з субстратом гіфами серцевини.
Водоростевий
компонент лишайника (фікобіонт) представлений синьо-зеленими, зeлeними,
жовіто-зеленими і бурими водоростями. Представники 28 родів цих відділів
водоростей вступають у симбіоз з грибами. Найпоширеніший фікобіонт лишайників –
зелена водорість требуксія, з інших зелених – хлорела, хлорокок. Із
синьо-зелених водоростей найбільш звичайними в таломі лишайників є носток
і глеокапса.
Більшість
цих водоростей у природі живе самостійно, але деякі зустрічаються лише в
лишайниках (требуксія, кокомікс), у вільному стані вони поки-що не виявлені.
Водорості в слані лишайшіка дуже змінюють свій зовнішній вигляд. Особливо це
стосується нитчастих водоростей, які в лишайнику розпадаються до окремих клітин
і змінюються до невпізнанності. В лишайнику водорості стають більш стійкими до
високих температур, можуть переносити тривале висушування. При культивуванні їх
на штучних середовищах (окремо від грибів) набувають вигляд, характерний для
форм, що живуть у природі.
Грибний
компонент лишайника представлений деякими видами сумчастих грибів і
базидіоміцетів. Кожний з компонентів лишайника зберігає властивий йому спосіб
розмноження. Сумчасті гриби утворюють на поверхні талому лишайника апотеції або
перитєції з сумками та сучкоспорами. Спори розносяться вітром, за сприятливих
умов проростають у гіфи, які, зустрівши клітини відповідного виду водорості,
обплітають їх і утворюють лишайник; якщо гіфи гриба не зустрінуть водорості,
вони гинуть. Водорість може існувати самостійно.
Слань
лишайників різноманітна за формою, розмірами, будовою, забарвленням. Колір
слані зумовлений наявністю пігментів у оболонках гіф і плодових тілах грибів.
Розрізняють п'ять груп пігментів: зелені, сині, фіолетові, червоні і коричневі.
Пігменти утворюються тільки на світлі. Чим більше світла в місці існування
лишайників, тим яскравіше вони мають забарвлення. За морфологічними ознаками
лишайники поділяють на коркові, листуваті і кущисті.
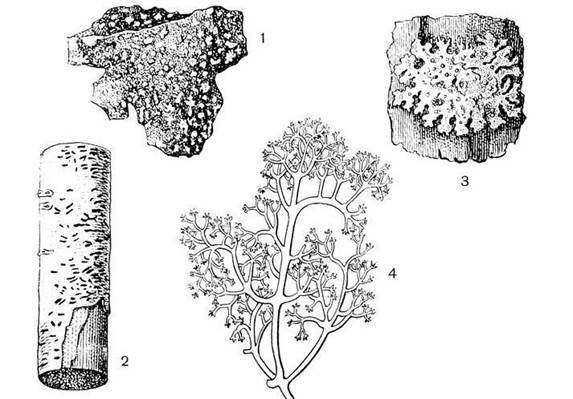
Рис. 18.10.
Морфологічні форми слані лишайника: 1. Накипний на камінні, 2. Накипний на корі
дерев, 3. Листуватий, 4. Кущистий
У коркових,
або накипних, лишайників слань має вигляд забарвленої кірки або нальоту, що
приростає дуже щільно до субстрату. Товщина кірки різна – від ледве помітного
накипу або порошкоподібного нальоту до 0.5 см, діаметр – від кількох міліметрів
до 20-30 см. Ростуть накипні лишайники на поверхні ґрунтів, гірських порід,
корі дерев і кущів, оголеній деревині, що гниє. До цієї групи лишайників
належить багато видів (близько 80%), що зустрічаються в різних умовах.
Листуваті
лишайники мають форму пластинок різного забарвлення, розташованих горизонтально
на субстраті (пармелія). Звичайно пластинки округлі, 10-20 см у діаметрі.
Характерною особливістю листуватих лишайників є неоднакове забарвлення і будова
верхньої і нижньої поверхні слані. У більшості з них на нижній частині слані
утворюються органи прикріплення до субстрату – ризоїди, які складаються із
зібраних у нитки гіф. Ростуть вони на поверхні ґрунту, серед мохів, на корі
дерев. Листуваті лишайники порівняно з накипними є більш високоорганізованими.
У кущистих
лишайників слань стеблоподібної форми, прикріплюється до субстрату невеликими
ділянками нижньої частини, а верхня частина розгалужена і піднята над поверхнею
або звисає з дерев – "бородаті лишайники". За рівнем організації
кущисті лишайники – вищий етап розвитку слані, їхня слань буває різних
розмірів: від кількох міліметрів до 30-50 см, Бородаті лишайники можуть
досягати 7-8 м (уснеа). До кущистих лишайників відносять ягель, цетрарію,
алекторію, нейропогон, евернію та ін.
Живуть
лишайники довго, сотні та навіть тисячі років. Вік знайденого в Гренландії
лишайника приблизно 4500 років.
Розмножуються
лишайники в основному вегетативно – частинами слані, які не є спеціалізованими
органами вегетативного розмноження. Крім того, розмноження проходить ізидіями
(виростами слані), а також соредіями (невеличкими утворами, які складаються з
клітин водоростей, обплетених гіфами грибів). Соредії та ізидії – особливі
"органи" розмноження лишайників як комплексних організмів. Соредії
утворюються всередині слані в гонідіальному шарі листуватих та кущистих
лишайників. Сформовані соредії виштовхуються із слані назовні, підхоплюються і
розносяться вітром. Скупчення соредій на поверхні лишайника називається сораль.
За сприятливих умов вони проростають у нових місцях і утворюють нові лишайники.
Соредіями розмножується близько 30% лишайників.
Ізидії –
вирости верхньої кірки слані. Мають вигляд зернин: циліндричних або
коралоподібних виростів або маленьких листочків. Всередині цих виростів
знаходяться клітини водоростей, оточених гіфами гриба. На відміну від соредій
ізидії не висипаються на поверхню слані, а разом з його шматочками відламуються
тваринами чи людиною і в сприятливих умовах розростаються у лишайник.
Розмноження ізидіями зустрічається у 15% лишайників.
Живлення
лишайників здійснюється за рахунок фотосинтезу у клітинах водоростей.
Синтезовані при цьому органічні речовини використовуються грибом. Дихання,
поглинання води і мінеральних солей забезпечує грибний компонент (мікобіонт)
слані лишайника. Активність процесу фотосинтезу, дихання, поглинання води і
мінеральних солей залежить від освітленості, температури, вологості.
Інтенсивність фотосинтезу у лишайників при оптимальних умовах значно нижча, ніж
у автотрофних рослин. Проте органічних речовин утворюється достатньо, щоб
забезпечити нормальну життєдіяльність лишайників.
Лишайники невибагливі до умов
середовища і характеризуються високою стійкістю до впливу несприятливих
факторів. Вони можуть рости у найрізноманітніших умовах освітлення і вологості,
легко переносять трипалі періоди без води, різкі коливання температури, але
по-різному реагують на забруднення повітря. Деякі з них не витримують навіть
малого забруднення повітря і гинуть, інші – живуть в населених пунктах, в тому
числі і промислових містах. Знаючи цю особливість лишайників, їх можна
використовувати як біоіндикатори дія оцінки чистоти повітря.
Характерна біологічна
особливість лишайників – утворення так званих лишайникових кислот, які
відкладаються на поверхні гіф у вигляді кристалів, паличок, зерняток і т.д.
Ними зумовлений колір лишайників. Відомо до 150 лишайникових кислот, які, крім
лишайників, ніде не зустрічаються. Біологічне значення їх ще не вивчене. Деякі
з них мають антибіотичні або токсичні властивості і, очевидно, виконують
захисну функцію.
У зв'язку з
широким розповсюдженням лишайники відіграють важливу роль у природі як
продуценти біомаси. Селячись на гірських породах, вони сприяють вивітрюванню
їх, а після відмирання утворюють невелику кількість гумусу, на якому можуть
оселятися інші рослини. Ось чому їх називають "піонерами
рослинності".
Лишайники
служать укриттям і їжею для багатьох безхребетних тварин. Ними живляться і
деякі крупні хребетні, наприклад, олені.
Лишайники
широко використовує людина в своїй господарській діяльності. Передусім, це
цінний корм для північних оленів (ягель). Деякі з них використовує в їжу людина
(цетрарія ісландська, умбілікарія їстівна). В їжу людина використовує також
види роду аспіцилія, відомі під назвою "манна небесна". Із лишайників
отримують спирт (цетрарія ісландська, деякі види кладоній), лакмус (леканора,
рочела), фарби (охролехія, деякі види рочел), їх використовутоть як сировину
для парфумерної промисловості (евернія сливова), в медицині для виготовлення
ліків (цетрарія, леканора, лобарія та ін.).

